 2024-12-23
2024-12-23 1365
1365 来源: Advanced Science
来源: Advanced Science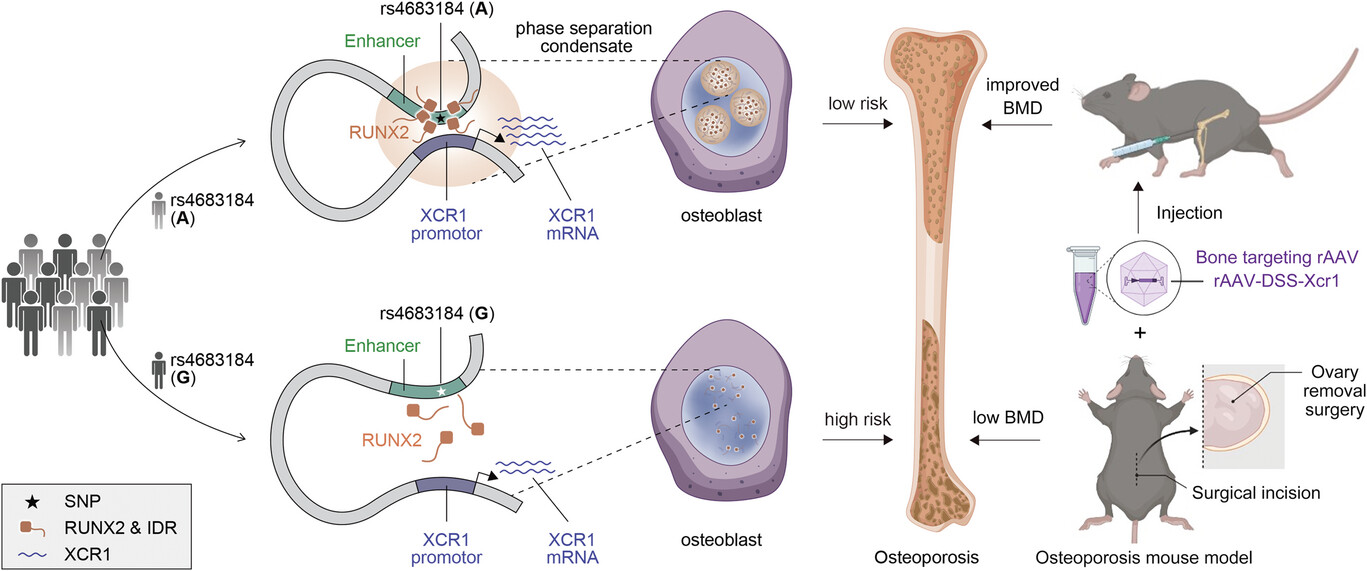

Advanced Science丨RUNX2 相分离介导骨质疏松易感变异体与 XCR1 之间的长程调控,促进成骨细胞分化
骨质疏松症是老年人不可避免的退行性骨骼疾病,其特征是骨密度 (BMD)低,遗传性高[1-4]。先前的全基因组关联研究 (GWASs) 已成功鉴定出与 BMD 和骨质疏松症相关的多个位点[5]。一些GWAS基因已被用作治疗骨质疏松症的药物靶点,并已成功进入临床试验或已上市,如钙传感受体、Dickkopf相关蛋白1、硬化蛋白和核因子κB配体的受体激活剂[4、6]。然而,GWASs 鉴定的大多数位点位于非编码区域,这对 GWAS 发现转化为临床应用提出了很大的挑战。
G 蛋白偶联受体 (GPCRs) 是调节成骨细胞分化和骨重塑的重要且必要,GPCR 家族相关基因可能是临床治疗骨质疏松症的关键靶点[7、8]。一般来说,具有遗传支持的药物靶点在临床试验中成功的可能性是其两倍[9]。具体来说,GWAS 已经鉴定了一些位于 X-C 基序趋化因子受体 1 (XCR1) 附近的非编码易感单核苷酸多态性 (SNP),这与 BMD 相关。XCR1,也称为 G 蛋白偶联受体 5 (GPR5),是 GPCR 家族的重要成员[10]。假设 XCR1 可能是骨质疏松症的新潜在靶点,但 GWAS SNP和 XCR1 之间的功能作用仍有待探索。
2024年12月20日,西安交通大学杨铁林,郭燕共同通讯在Advanced Science 在线发表题为“RUNX2 Phase Separation Mediates Long-Range Regulation Between Osteoporosis-Susceptibility Variant and XCR1 to Promote Osteoblast Differentiation”的研究论文。该研究首次将非编码SNP与相分离联系起来,为研究复杂疾病易感性的远端染色质调控机制提供了新的视角,并为骨质疏松药物的开发和相应的转化研究找到了潜在靶点。

在本研究中,我们阐明了 GWAS 基因间 SNP (rs4683184) 作为增强子的功能基础,它改变了 RUNX 家族转录因子 2 (RUNX2) 的结合亲和力并导致差异 XCR1 表达和成骨细胞分化。重要的是,我们发现 RUNX2 相分离可以介导 rs4683184 和 XCR1 之间的长程调节。XCR1 可促进成骨细胞分化。携带 Xcr1 基因的骨靶向腺相关病毒 (AAV) 可以改善骨质疏松症小鼠的骨形成并增加 BMD。总之,我们的研究结果不仅阐明了转录因子和非编码 SNP 通过相分离影响疾病易感性的新调控机制,而且为骨质疏松症药物的开发提供了新的靶点。
虽然 GHAS 已经确定了许多与骨质疏松症相关的基因座,但潜在的遗传调控机制和潜在的药物靶点仍有待探索。在我们的研究中,我们破译了一种新的调控机制,即 GWAS 基因间 SNP (rs4683184) 作为增强子影响 RUNX2 的结合亲和力,其相分离可以介导增强子和靶基因 XCR1 之间的长程染色质相互作用,导致 XCR1 表达和成骨细胞分化的变化。Xcr1 的骨靶向 AAV 可以改善骨质疏松症小鼠的骨形成并增加 BMD,表明 XCR1 可能是骨质疏松症的新易感基因(图1)。我们的研究为理解 TFs 和非编码增强子 SNP 的机制提供了新的视角,并为骨质疏松症药物的开发和相应的转化研究找到了潜在的靶点。
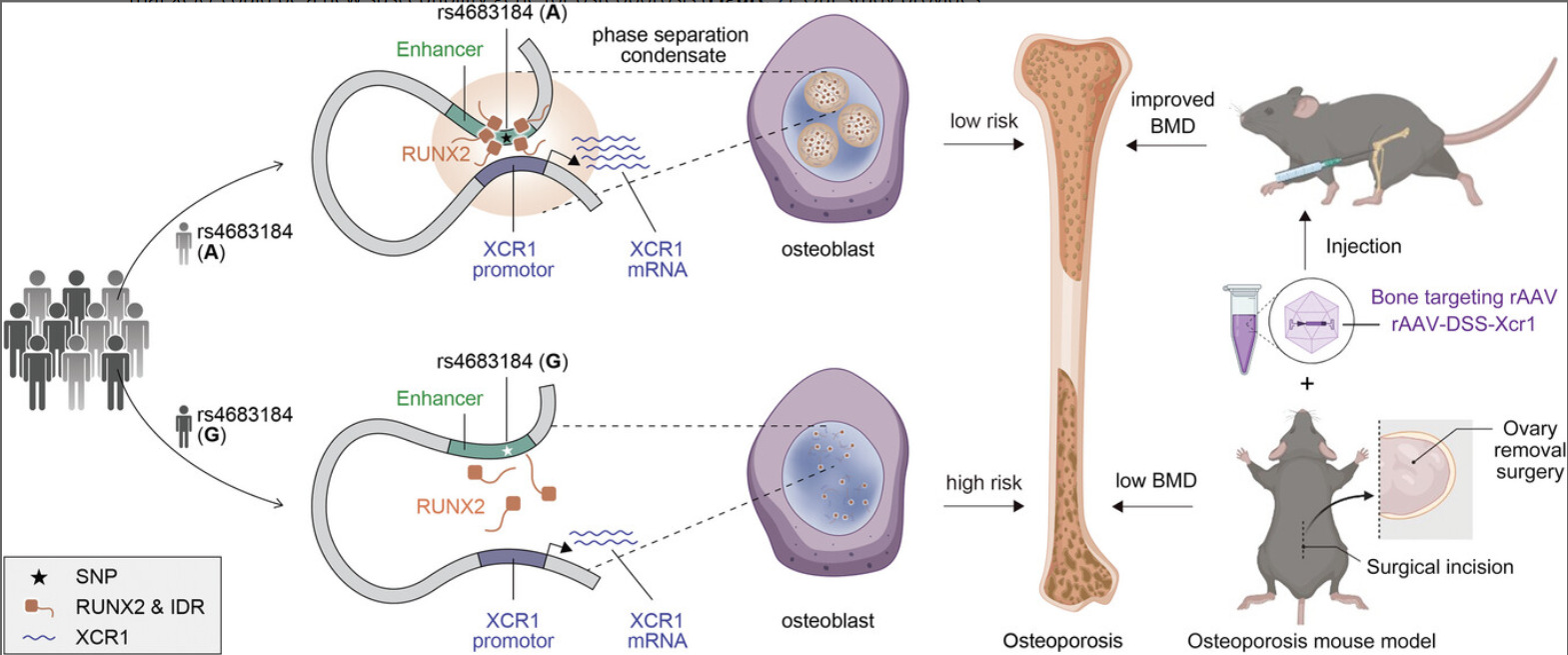
图1
综上所述,我们的研究揭示了 RUNX2 相分离介导非编码易感性 SNP rs4683184 与靶基因 XCR1 之间的长程相互作用,进而调节参与成骨细胞分化和骨质疏松症的 XCR1 的表达。这些结果不仅有助于我们从新的角度理解成骨细胞分化中的远程转录调控过程,而且阐明了转录因子和非编码 SNP 通过相分离影响疾病易感性的新机制。预计该策略将显着加速骨质疏松症新药的开发,为临床实践提供更有效的治疗选择。
原文出处:https://onlinelibrary.wiley.com/doi/10.1002/advs.202413561
图片来源网络,如有侵权请及时联系删除!

